
Суеверия голубя
Автор: B. F. Skinner
Перевод: Жесткова Е.Б.
Заявление о том, что подкрепление обусловлено реакцией, может означать только то, что оно следует за реакцией. Подкрепление может быть обусловлено механической связью, или опосредовано действиями другого живого организма; но можно предположить, что обусловленность имеет место исключительно вследствие временной связи, которая проявляется в последовательности и близости реакции и подкрепления. Когда мы представляем положение дел, которое, как мы знаем, является подкреплением при соблюдении определённого темпа и временных интервалов, мы всегда должны предположить, что имеет место обусловленность, даже несмотря на то, что, представляя положение дел, мы проигнорировали поведение организма. Простой эксперимент демонстрирует нам, что это действительно так.
Голубя приводят в стабильное состояние голодания, сокращая его вес на 75% от его исходного веса, когда он получал хорошее питание. Далее птицу помещают в экспериментальную клетку на несколько минут каждый день. Кормушка, прикрепленная к клетке, может быть поставлена на место, чтобы голубь мог из неё поклевать. Электромагнит и временное реле удерживают кормушку на месте в течение пяти секунд при каждом подкреплении.
Если теперь установить часы, чтобы подавать корм в кормушку через регулярные промежутки времени вне зависимости от поведения птицы, то, как правило, происходит оперантное обусловливание. В шести из восьми случаев реакции были настолько четко определены, что результат подсчета случаев полностью совпадал у двух наблюдателей. Одну птицу приучили вращаться по клетке против часовой стрелки, делая два или три оборота между подкреплениями. Другая неоднократно упиралась головой в один из верхних углов клетки. У третьей развилась реакция «подбрасывания», как будто она помещает голову под невидимую планку и многократно поднимает ее. У двух птиц развилось маятниковое движение головы и тела, при котором голова выдвигалась вперед и раскачивалась справа налево с резким движением, за которым следовало чуть более медленное возвращение в исходное положение. Тело, как правило, следовало за движением, и приходилось делать несколько шагов, когда движение было продолжительным. Ещё одна птица была обучена делать неполные клевательные или имитирующие чистку перьев движения, направленные в пол, но при этом не касающиеся его. Ни одна из этих реакций не проявилась с заметной силой в период адаптации к клетке или до тех пор, пока кормушка не стала появляться периодически. В других двух случаях условные реакции не были четко выражены.
Процесс обусловливания обычно очевиден. Птица демонстрирует какую-то реакцию в момент появления кормушки; в результате она стремится повторить эту реакцию. Если интервал перед следующим предоставлением кормушки не настолько большой, чтобы реакция исчезла, вероятно повторное «обусловливание». Это еще больше укрепляет реакцию, и повышает вероятность последующего подкрепления. Верно то, что некоторые реакции остаются без подкрепления, а некоторые подкрепления появляются ещё до появления реакции, однако в итоге развивается значительное состояние силы.
За исключением поворота против часовой стрелки, каждая реакция почти всегда повторялась в одной и той же части клетки и, как правило, была связана с ориентацией на какую-то особенность клетки. Эффект подкрепления заключался в том, чтобы приучить птицу реагировать на какой-то аспект окружающей среды, а не просто выполнять серию движений. между подкреплениями все реакции быстро повторялись – по пять или шесть раз за 15 секунд.
Эффект, по-видимому, зависит от скорости подкрепления. В целом можно ожидать, что чем короче интервал между подкреплениями, тем быстрее и заметнее наступает обусловливание. Одна из причин заключается в том, что поведение голубя становится более разнообразным по мере того, как проходит время после подкрепления. Сто фотографий, каждая из которых сделана спустя две секунды после того, как убрали кормушку, демонстрируют довольно однообразное поведение. Птица находилась бы в одной и той же части клетки, рядом с кормушкой и, возможно, ориентировалась бы на стенку, где была кормушка, или поворачивалась бы в ту или иную сторону. Сотня фотографий, сделанных спустя 10 секунд, напротив, зафиксировала бы птицу в разных частях клетки, реагирующую на разные аспекты окружающей среды. Поэтому, чем быстрее появляется второе подкрепление, тем больше вероятность того, что вторая подкрепленная реакция будет похожа на первую, а также вероятность того, что обе реакции будут выражены в одной из нескольких стандартных форм. В случае очень короткого интервала, который станет ограничением, можно ожидать, что голубь повернёт голову в сторону отверстия, в котором исчезли припасы.
Ещё одна причина большей эффективности коротких интервалов заключается в
том, что чем длиннее интервал, тем больше количество промежуточных реакций без подкрепления. В результате исчезновение реакции отменяет эффект случайного подкрепления.
Согласно этой интерпретации, эффективность интервала будет зависеть от скорости обусловливания и скорости исчезновения, и, следовательно, будет варьироваться в зависимости от скорости, а также, предположительно, от биологического вида. Пятнадцать секунд — очень эффективный интервал при указанной выше скорости. Интервал длительностью одной минуты гораздо менее эффективен. Однако, когда реакция уже установлена, длительность интервала может быть увеличена. В одном случае она была увеличена до двух минут, и высокая скорость реакции сохранялась без каких-либо признаков ослабления. В другом случае реакции наблюдались в течение многих часов с интервалом в одну минуту между подкреплениями.
В последнем случае реакция демонстрировала заметный сдвиг топографии. Он начинался как резкое движение головы из центра влево. Это движение становилось все более энергичным, и в конце концов, птица поворачивалась в том же направлении и делала шаг или два. Спустя много часов реакция с помощью шагов стала преобладающей. Птица совершала хорошо выраженный прыгающий шаг с правой лапки на левую, одновременно поворачивая голову и туловище влево, как и раньше.
Когда реакция, выраженная в шаге, стала проявленной, стало возможным сделать механическую запись, поместив птицу на большой табурет, непосредственно соединенный с маленьким табуретом, и с каждым шагом птицы происходил тонкий электрический контакт. Наблюдая за птицей и прислушиваясь к звуку записи, можно было убедиться в том, что ведется достаточно достоверная запись. Птица могла слышать звук диктофона при каждом шаге, но это, конечно, никак не коррелировало с кормлением. Запись, полученная при предоставлении пищи каждую минуту, во всех отношениях напоминает характерную кривую для голубя при периодическом подкреплении стандартной выбранной реакции. Развивается хорошо заметная временная дискриминация. Птица не реагирует сразу после еды, но по истечении 10, 15 или даже 20 секунд она начинает быстро реагировать и продолжает это делать до тех пор, пока не будет получено подкрепление.
Когда часы были выключены, и пища не предоставлялась, можно было зафиксировать «угасание» реакции. Птица продолжала реагировать характерными прыжками из стороны в сторону. Было зарегистрировано более 10 000 реакций, прежде чем «угасание» достигло той точки, когда в течение 10 или 15 минут было несколько
реакций, если они вообще были. Когда часы были снова запущены, периодическое предоставление пищи (по-прежнему без какой-либо связи с поведением птицы) привело к появлению типичной кривой для восстановления после периодического подкрепления, показанной на рис. l). До начала этой кривой пластинка была практически горизонтальной в течение 20 минут. Первое подкрепление имело незначительный эффект, а второе — более значительный. Наблюдалось плавное ускорение темпа в положительную сторону, когда птица возвращалась к преобладающей скорости реакции, когда подкрепление предоставлялось ей каждую минуту.
Когда реакция была снова погашена и периодическое предоставление пищи возобновилось, была получена другая реакция. Эта реакция проявлялась в ходьбе, при которой птица перемещалась по клетке. Реакция, выраженная в прыжках из стороны в сторону больше не появлялась и, конечно, не могла быть получена намеренно, если не ставить подкрепление в зависимость от поведения.
Можно сказать, что этот эксперимент демонстрирует своего рода суеверие. Птица ведет себя так, как будто между ее поведением и подачей корма существует причинно-следственная связь, хотя такие отношения отсутствуют. В человеческом поведении существует множество аналогий. Хорошим примером являются ритуалы, направленные на привлечение удачи в
картах. Нескольких случайных связей между ритуалом и благоприятными последствиями достаточно, чтобы установить и поддерживать поведение, несмотря на множество случаев без подкрепления. Ещё один пример можно привести из игры в боулинг: игрок, выпуская шар на дорожку, продолжает вести себя так, словно он управляет шаром, выворачивая руку и плечо. Разумеется, такое поведение не оказывает никакого особого влияния на удачу или на шар, катящийся в заданном направлении по дорожке, также как в данном случае корм появлялся бы так же часто, если бы голубь не делал ничего — или, точнее, делал что-то другое.
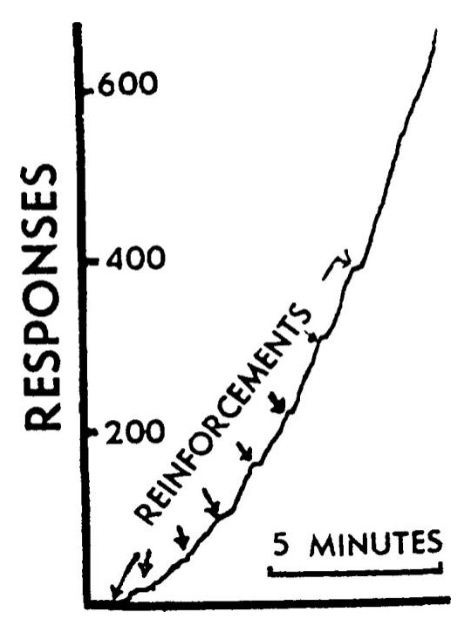
Рис.1 «Восстановление» суеверной реакции после ее угасания. (Реакция, выраженная в прыжках справа налево была тщательно погашена непосредственно перед тем, как была сделана запись. Стрелки указывают на автоматическое предоставление пищи с интервалом в одну минуту без учета поведения голубя).
Возможно, не совсем корректно говорить, что обусловленное поведение было установлено без каких-либо заранее определенных условий. Мы апеллировали к единой последовательности реакций в поведении голубя, чтобы получить чистую условную реакцию в целом. Когда мы устанавливаем часы так, чтобы давать голубю корм каждые 15 секунд, мы фактически основываем наше подкрепление на ограниченном наборе реакций, которые часто возникают спустя 15 секунд после подкрепления. Когда реакция усиливается (а это может произойти в результате одного подкрепления), установка часов подразумевает еще более ограниченные условия. Нечто подобное можно сказать и об игроке. Не совсем верным будет утверждение о том, что нет никакой связи между поворотами и разворотами его туловища и траекторией, пройденной шаром до дальнего конца дорожки. Связь была установлена до того, как шар выскочил из руки игрока, но поскольку и траектория шара, и поведение игрока определены, — некоторая связь сохраняется. Последующее поведение игрока может не оказывать никакого влияния на шар, но поведение шара влияет на игрока. Эта зависимость, хоть и не является идеальной, будет достаточной для поддержания поведения в силе. Конкретная форма поведения, принятая игроком, вызвана индукцией, вызванной реакциями, в которых имеет место фактический контакт с шаром.Очевидно, что это движение соответствует изменению направления движения шара. Но это не отменяет ценности сравнения, поскольку нас интересует не то, какая реакция выбрана, а то, почему она сохраняет свою силу. В ритуалах привлечения удачи индуктивное усиление конкретной формы поведения, как правило, отсутствует. Поведение голубя в этом эксперименте относится к последнему типу, о чем свидетельствует разнообразие реакций, полученных при исследовании поведения разных голубей. Вопрос о том, существует ли у голубя необусловленное поведение, соответствующее определенному воздействию на окружающую среду, изучается по сей день.
Полученные результаты проясняют случайное поведение, наблюдаемое в экспериментах, в которых часто представляется дискриминационный стимул. Такой стимул имеет подкрепляющую ценность и может вызвать поведение, в котором можно увидеть взаимосвязь с внешними факторами. Голубь часто вырабатывает определенную реакцию, например, поворачивается, крутится, клюет вблизи места нахождения дискриминационного стимула, хлопает крыльями и т.д. В большинстве работ, проведенных до настоящего времени в этой области, интервал между дискриминационными стимулами составлял одну минуту, и многие из этих реакций, обусловленных внешними факторами, были кратковременными. Не вызывает сомнений тот факт, что появление этих реакций является результатом случайных корреляций с предоставлением стимула.